
Виталий Шевченко
«Природа» №5, 2019
 Об автореВиталий Андреевич Шевченко — кандидат биологических наук, научный сотрудник лаборатории перспективных исследований мембранных белков Московского физико-технического института, Института комплексных систем Исследовательского центра Юлиха и соискатель степени PhD Института кристаллографии Рейнско-Вестфальского технического университета Ахена. Область научных интересов — поиск новых оптогенетических инструментов, их описание и разработка (инжиниринг). Победитель конкурса «Био/мол/текст — 2017». |
До недавнего времени исследования мозга могли обрисовать его работу лишь «крупными мазками». Современные технологии, такие как томография, позволяют «разглядывать» мозг с высоким пространственным разрешением, причем происходит это неинвазивно, т.е. нетравматично. Относительно недавно был сделан еще один шаг, позволяющий не только наблюдать, но и управлять нервными импульсами, — разработаны оптогенетические технологии. Оптогенетика — это способ управления определенными клетками с помощью света. Чаще всего речь идет о нейронах и мышечных клетках — так называемых электровозбудимых клетках. Как это работает? Для нормального существования клетка старается поддерживать определенные условия внутри и снаружи себя. По разные стороны мембраны клетки находятся ионы, но в различных концентрациях. В частности, внутри клетки преобладают ионы калия, а снаружи — натрия и хлора. Причем, соотношение заряженных ионов таково, что клетка внутри заряжена более отрицательно (имеет отрицательный мембранный потенциал, равный примерно −70 мВ). Теперь, если мы каким-либо образом извне направим положительные заряды внутрь клетки, произойдет ее деполяризация, и клетка возбудится (возникнет потенциал действия). У нейронов в мембране находятся свои белки, которые реагируют на изменение потенциала. Именно они отвечают за распространение нервного импульса по цепочке нейронов.
В природе есть различные организмы, жизнь которых зависит от наличия света. Поглощая фотоны, естественные белки в этих организмах могут переносить различные ионы через мембрану клеток (например, протоны или ионы натрия). Если в нейрон или мышечную клетку внедрить такой белок, переносящий положительные ионы внутрь, и посветить на эту клетку, то мы искусственно запустим нервный импульс. Мы также можем и заблокировать прохождение импульса: для этого нужно, напротив, еще сильнее уменьшить мембранный потенциал — так, чтобы собственные клеточные белки не справились с повышением потенциала до нужного порога. Такое уменьшение называется гиперполяризацией. Этого можно добиться, выкачивая положительные ионы из клетки или закачивая в нее отрицательные.
Обобщая, можно сказать, что оптогенетика находит белки-транспортеры, реагирующие на свет, пытается доставить эти белки в нужные клетки и создает решения, как же посветить на эти клетки светом (это особенно сложно, когда хочется светить на клетки глубоко в мозге). Причем эта система модульная, ее составные компоненты можно заменять.
Доступно и подробно о том, как работает оптогенетика как таковая, можно прочитать в статье В. Попова «Светлая голова», опубликованной на сайте «Биомолекула». Здесь же мы сфокусируемся на работе белков — молекулярных инструментах оптогенетики.
Светочувствительные белки
В природе разнообразие белков-транспортеров (опсинов) действительно велико. Ими обзавелись различные бактерии, археи, простейшие и даже вирусы. Кому-то такие белки нужны, чтобы вырабатывать энергию. Например, солеустойчивые (галофильные) археи Halobacterium halobium пользуются своей мембраной буквально как солнечной батарейкой. В начале 1970-х годов у этих микроорганизмов был обнаружен белок бактериородопсин, который под действием света выкачивает протоны наружу клетки, и созданная таким образом разность потенциалов на мембране используется клеткой для синтеза АТФ [1, 2]. Вскоре после открытия бактериородопсина были обнаружены еще два сенсорных родопсина (известные также как медленные родопсины) в другой галофильной архее H. salinarum [3, 4]. Кроме того, в ней был найден еще и четвертый родопсин (галородопсин), который функционирует как хлоридный насос [5, 6].
Другие археи-экстремалы, живущие в щелочных водоемах, чтобы как-то существовать, тоже завели у себя специальный белок — ксенородопсин, который при поглощении фотона закачивает внутрь клетки протон, поддерживая кислотно-щелочной баланс в цитоплазме. В 2013 г. японские ученые обнаружили у бактерий Krokinobacter eikastus новый микробный родопсин, выкачивающий натрий из клетки, — KR2 (от K. eikastus rhodopsin 2) [7, 8].
У некоторых простейших светочувствительные белки «настроены» на синий / фиолетовый спектр. Каскадом химических реакций они связаны с работой жгутиков клеток, что позволяет организмам уплывать от губительного действия коротковолнового (фиолетового-ультрафиолетового) излучения. У человека тоже есть светочувствительные белки: к примеру, в палочках сетчатки глаза находятся родопсины, которые отвечают за восприятие света [9].
Классические оптогенетические инструменты — галородопсин, археародопсин и канальный родопсин (рис. 1) [10]. Галородопсин и археародопсин используются для деактивации нейронов, они блокируют передачу нервного импульса. Под действием света галородопсин переносит отрицательно заряженные ионы хлора внутрь клетки, тем самым вызывая гиперполяризацию нейрона. Археародопсин выкачивает протоны из клетки, что, естественно, также гиперполяризует нейрон. И галородопсин, и археародопсин — это так называемые насосы, помпы. Если вокруг есть нужный им ион (а он есть почти всегда), то, поглотив квант света, эти белки активно перенесут ион с одной стороны мембраны на другую.

Канальный родопсин в настоящее время широко используют для активации электровозбудимых клеток. Он долгое время оставался единственным белком, способным на это. Канальный родопсин позволяет положительным ионам перетекать из области с их большей концентрацией в область с меньшей. Из внеклеточного пространства внутрь клетки идет поток ионов натрия, и нейрон возбуждается.
Несмотря на то, что три классических белка-транспортера в принципе решают задачи активации и деактивации электровозбудимых клеток, у каждого из них есть свои преимущества и недостатки. Например, галородопсин недостаточно быстро переносит ионы хлора, а археародопсин в своей работе использует протоны и меняет кислотность среды вокруг нейрона, отчего клетка может начать чувствовать себя неважно. Канальный родопсин — замечательный инструмент, но переносит ионы только пассивно: ему нужна разность концентраций ионов, чтобы все работало. Кроме этого, канальный родопсин не избирателен к ионам и эффективен, только если его освещать синим или фиолетовым светом, а такое коротковолновое излучение очень неглубоко проникает в ткани (рис. 2). Это ограничивает возможность применения белка, если интересующий нас участок мозга или мышцы находится глубоко. Именно поэтому во множестве лабораторий не прекращается поиск, разработка и оптимизация оптогенетических инструментов нового поколения.
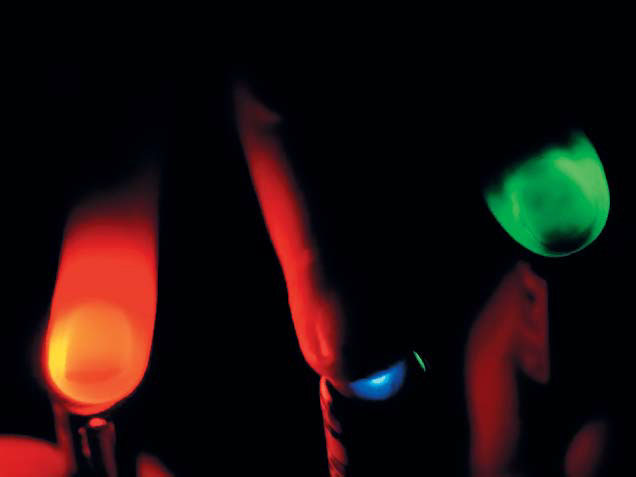
Оптогенетические инструменты будущего
В последнее время существенно изменили и подстроили параметры работы популярного канального родопсина: с помощью направленного введения мутаций можно получить его аналоги, более быстрые и специфичные к ионам кальция, и с оптимумом работы, сдвинутым в более длинноволновую часть спектра.
Исследование баз данных белков и организмов позволяет отыскивать новые белки-транспортеры, пригодные для нужд оптогенетики. Каждая из этих находок ценна по-своему. Открытые анионные канальные родопсины [11] позволяют деактивировать нейроны, создавая поток отрицательных зарядов внутрь клетки. Химерный белок, состоящий из фоточувствительной части и вирусного калиевого канала [12], может деактивировать нейроны, пропуская ионы калия из цитоплазмы во внеклеточное пространство.
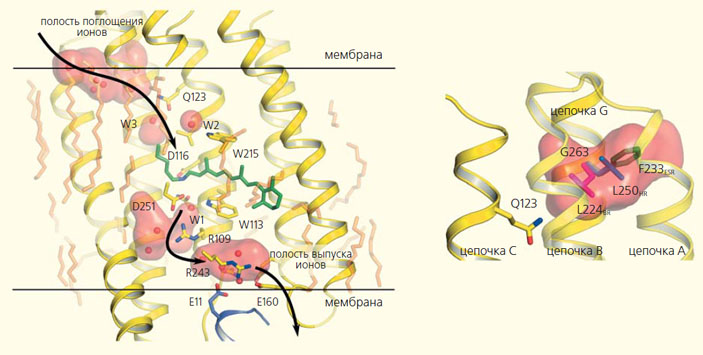
В 2015 г. нам с коллегами удалось закристаллизовать белок KR2, открытый несколько лет назад японскими учеными [7, 8], определить его структуру в различных функциональных состояниях и разобраться в механизмах и условиях работы этого натриевого насоса (рис. 3) [13]. Оказалось, что у KR2 есть полость, отвечающая за избирательность иона, который будет перенесен. И внеся в белок мутации, затрагивающие эту полость, нам удалось получить модификацию белка, которая прокачивает ионы калия [8, 13]. А ведь именно на паре ионов натрия / калия работают собственные клеточные белки, ответственные за прохождение нервного импульса. Таким образом, на основе полученных данных мы смогли создать более естественный, физиологический оптогенетический инструмент. Это пример того, как человек, используя свои знания и воображение, может подстраивать под собственные нужды то, что придумала природа.
Наша группа включает в себя тех, кто может произвести белок в большом количестве, кто имеет большой опыт в кристаллизации мембранных белков (а это очень непростая задача) и кто умеет определять структуру белка, получив данные рентгеновской дифракции. Затем мы анализируем полученную информацию и проводим дальнейшие эксперименты с модификациями белка. Новые функции мутантных форм исследуем электрофизиологически, методом локальной фиксации потенциала. Финальным аккордом мы стараемся продемонстрировать применимость изученного белка в нервных клетках.
В 2017 г. мы фактически бросили вызов монополисту — канальному родопсину, открыв протонный насос, переносящий протоны внутрь клетки [14]. Чтобы его найти, было исследовано множество таких родопсинов, чья функция была еще неизвестна. Изучая разные микроорганизмы, мы старались найти те, которые живут в каких-то экзотических условиях. Пришлось перебрать с десяток кандидатов, прежде чем удалось найти тот, который давал бы надежду на дальнейшее практическое применение. Наш протонный насос относится к семейству ксенородопсинов, а обнаружен он был в крошечных археях (наногалоархеях), которые обитают в щелочных водоемах. Мы считаем, что эти организмы используют белок для поддержания гомеостаза, преодолевая негативное действие окружающей среды. Ксенородопсины эффективно работают, поглощая красный свет, что позволяет использовать их глубоко в тканях. Поэтому мы смело утверждаем, что открытые белки могут составить достойную конкуренцию канальному родопсину.
Ксенородопсины переносят протон достаточно быстро, и уже через 2,5 мс готовы перенести следующий протон. Фактически это означает, что белок может возбуждать человеческие нейроны с максимальной скоростью. Ксенородопсины способны занять нишу работы с мышечными клетками. Дело в том, что мышечные клетки очень чувствительны к транспорту кальция, поэтому при работе с мышцами хотелось бы избежать его переноса, чего не может обеспечить канальный родопсин, так как он неселективен (см. рис. 1). А ксенородопсины могут.
Как и в случае с натриевым насосом, нам удалось определить структуру ксенородопсина (рис. 4, слева) и понять механизм переноса протона. Многие исследователи, которые работают в области фотоактивных белков, были очень удивлены тому, что такой белок мог найтись в природе. Широко распространено мнение, что перенос протонов через мембрану подобных микроорганизмов в первую очередь обеспечивает клетку энергией. А поведение ксенородопосина не вписывалось в эту картину. В связи с этим мы специально показали на модельных клетках HEK293, что есть ток внутрь (см. рис. 4, справа сверху), причем направление тока оставалось тем же, даже если к клетке приложить удерживающее напряжение. Ксенородопсины способны насильно вкачивать протоны в цитоплазму. В этой же работе мы поставили восклицательный знак, на деле показав, что белок может возбуждать нейроны с высокой частотой (см. рис. 4, справа снизу).

Наконец, в апреле текущего года мы подробно описали, как работает натриевый насос KR2 в физиологических условиях [15]. Мы — это международная группа ученых из Центра исследований молекулярных механизмов старения и возрастных заболеваний Московского физико-технического института (Россия), Института структурной биологии Гренобльского университета и Европейского ускорительного комплекса в Гренобле (Франция), Института комплексных систем Исследовательского центра Юлиха (Германия), Института кристаллографии Рейнско-Вестфальского технического университета и Института Макса Планка (Германия).
В результате работы выяснилось, что для функционирования белка ему необходимо собираться в пентамеры — структуры, состоящие из пяти субъединиц белка (рис. 5). Более того, исходя из данных спектроскопических измерений, все субъединицы находятся в разных состояниях, что весьма необычно. Необходимость в таком исследовании возникла потому, что несколько групп ученых опубликовали данные, в которых кристаллические структуры важного для прикладных исследований белка имели различия, которые нужно было объяснить. Оказалось, что многообразие структур KR2 связано с тем, что авторы публикаций изучали новый белок не в совсем идентичных условиях. Напомню, что микроорганизм, синтезирующий белок с уникальными свойствами натриевого насоса, обитает в океане, где концентрация NaCl составляет примерно 500 мМ и pH — около 8. Только в этих условиях KR2 качает ионы натрия, формируя при этом пентамеры в мембране клеток, а при низком pH этот белок действует как протонный насос. Разнообразные «ложные» структуры белка, вероятно, либо оказались артефактами кристаллизации, либо изучались в ненадлежащих условиях. Смоделировав физиологические условия функционирования KR2, мы в результате описали его подлинную структуру. Понимание механизма работы родопсина KR2 важно не только для фундаментальных исследований работы нервной системы живых организмов, но и для медицинской практики — для дальнейшего рационального мутагенеза и создания новых инструментов оптогенетики.

***
Исследования мозга и минимально инвазивная терапия заболеваний, связанных с нервной системой, продолжают и продолжат быть актуальными в течение XXI в. Оптогенетика и ее инструменты — молекулярные машины и белки-транспортеры — дадут возможность улучшить качество жизни людей с перманентными болевыми синдромами, нейродегенеративными возрастными заболеваниями, помогут частично восстановить зрение [16, 17] и слух, а возможно, и мышечные дисфункции. Фундаментальные исследования, поиск и рациональная модификация белков будут лежать в основе успеха оптогенетических подходов в терапии.
С момента публикации работы о ксенородопсинах были предприняты попытки использовать его в оптогенетических кохлеарных имплантах — устройствах, компенсирующих потерю слуха [18, 19]. Поскольку ксенородопсин способен возбуждать нервные клетки с очень высокой частотой (она скорее определяется уже не свойствами белка, а скоростью возбуждения и релаксации нейронов), можно управлять нейронами улитки уха с высоким временным разрешением. Данная техника может составить конкуренцию электронным имплантам, у которых низкое пространственное разрешение возбуждаемых нейронов. Безусловно, использование оптогенетических инструментов в медицинских приложениях не за горами. Однако требуется решить важный вопрос, связанный с допустимостью применения генной терапии, когда для помощи пациенту потребуется вмешательство в его клетки на генетическом уровне.
Работа выполнена при поддержке Федеральной целевой программы «Исследования и разработки по приоритетным направлениям развития научного-технологического комплекса» (проекты 14.587.21.0011 323, RFMEFI58715X0011), а также Российским фондом фундаментальных исследований (проект 19-52-15017).
Литература
1. Oesterhelt D., Stoeckenius W. Rhodopsin-like protein from the purple membrane of Halobacterium halobium // Nat. New Biol. 1971; 233: 149–152. DOI: 10.1038/newbio233149a0.
2. Racker E., Stoeckenius W. Reconstitution of purple membrane vesicles catalyzing light-driven proton uptake and adenosine triphosphate formation // J. Biol. Chem. 1974; 249: 662–663.
3. Hildebrand E., Dencher N. Two photosystems controlling behavioural responses of Halobacterium halobium // Nature. 1975; 257: 46–48. DOI: 10.1038/257046a0.
4. Takahashi T., Tomioka H., Kamo N., Kobatake Y. A photosystem other than PS370 also mediates the negative phototaxis of Halobacterium halobium // FEMS Microbiol. Lett. 1985; 28: 161–164. DOI: 10.1111/j.1574-6968.1985.tb00784.x.
5. Matsuno-Yagi A., Mukohata Y. Two possible roles of bacteriorhodopsin; a comparative study of strains of Halobacterium halobium differing in pigmentation // Biochem. Biophys. Res. Commun. 1977; 78(1): 237–243. DOI: 10.1016/0006-291X(77)91245-1.
6. Schobert B., Lanyi J. K. Halorhodopsin is a light-driven chloride pump // J. Biol. Chem. 1982; 257(17): 10306–10313.
7. Inoue K., Ono H., Abe-Yoshizumi R. et al. A light-driven sodium ion pump in marine bacteria // Nat. Commun. 2013; 4: 1678. DOI: 10.1038/ncomms2689.
8. Kato H. E., Inoue K., Abe-Yoshizumi R. et. al. Structural basis for Na+ transport mechanism by a light-driven Na+ pump // Nature. 2015; 521, 48–53; DOI: 10.1038/nature14322.
9. Fotiadis D., Liang Y., Filipek S. et al. Atomic-force microscopy: Rhodopsin dimers in native disc membranes // Nature. 2003. 421: 127–128. DOI: 10.1038/421127a.
10. Sizemore R. J., Seeger-Armbruster S., Hughes S. M., Parr-Brownlie L. C. Viral vector-based tools advance knowledge of basal ganglia anatomy and physiology // J. Neurophysiol. 2016; 115(3): 2124–2146. DOI: 10.1152/jn.01131.2015.
11. Govorunova E. G., Sineshchekov O. A., Janz R. et. al. Natural light-gated anion channels: A family of microbial rhodopsins for advanced optogenetics // Science. 2015; 349(6248): 647–650. DOI: 10.1126/science.aaa7484.
12. Cosentino C., Alberio L., Gazzarrini S. et al. Engineering of a light-gated potassium channel // Science. 2015; 348(6225): 707–710. DOI: 10.1126/science.aaa2787.
13. Gushchin I., Shevchenko V., Polovinkin V. et al. Crystal structure of a light-driven sodium pump // Nat. Struct. Mol. Biol. 2015; 22(5): 390–395. DOI: 10.1038/nsmb.3002.
14. Shevchenko V., Mager T., Kovalev K. et al. Inward H+ pump xenorhodopsin: Mechanism and alternative optogenetic approach // Sci. Adv. 2017. 3(9): e1603187. DOI: 10.1126/sciadv.1603187.
15. Kovalev K., Polovinkin V., Gushchin I. et al. Structure and mechanisms of sodium-pumping KR2 rhodopsin // Sci. Adv. 2019; 5(4): eaav2671. DOI: 10.1126/sciadv.aav2671.
16. Suzuki K., Tsunekawa Y., Hernandez-Benitez R. et. al. In vivo genome editing via CRISPR/Cas9 mediated homology-independent targeted integration // Nature. 2016. 540, 144–149. DOI: 10.1038/nature20565.
17. Nandrot E. F., Silva K. E., Scelfo C., Finnemann S. C. Retinal pigment epithelial cells use a MerTK-dependent mechanism to limit the phagocytic particle binding activity of αvβ5 integrin // Biology of the Cell. 2012; 104(8): 326–341. DOI: 10.1111/boc.201100076.
18. Wrobel Ch., Dieter A., Huet A. et. al. Optogenetic stimulation of cochlear neurons activates the auditory pathway and restores auditory-driven behavior in deaf adult gerbils // Sci. Transl. Med. 2018; 10(448): eaao0540. DOI: 10.1126/scitranslmed.aao0540.
19. Dieter A., Duque-Afonso C. J., Rankovic V. et. al. Near physiological spectral selectivity of cochlear optogenetics // Nat. Commun. 2019; 10(1): 1962. DOI: 10.1038/s41467-019-09980-7.
Подробнее см.: Камкин А. Г., Киселева И. С., Ярыгин В. Н. Новый тип ионных каналов // Природа. 2002. № 3. С. 13–20. — Примеч. ред.
